наземные или реже пресноводные автотрофные растения, объединяемые в отдел мохообразных (См.
Мохообразные), включающий наиболее примитивные высшие растения. М. подразделяют на 3 класса: антоцеротовые, печёночные и лиственные, или листостебельные. Все они сравнительно просто организованные, многоклеточные, большей частью многолетние (реже - однолетние) растения высотой от 1 до 50
см с частичным обособлением ассимиляционной, водопроводящей и механической тканей; обладают единым циклом развития, различающимся в пределах отдельных групп некоторыми особенностями. Цикл развития М. сопровождается обособлением диплоидного
Спорофита (видоизменённого спорангия) на гаплоидном
Гаметофите. В результате половое и бесполое поколения развиваются совместно на одном растении, т. е.
Чередование поколений у М. носит несколько условный характер. Из споры у М. вырастает ветвистая, многоклеточная нитчатая или пластинчатая
Протонема, способная к фотосинтезу. Вследствие развития на протонеме многочисленных почек для М. характерны групповые формы роста (дерновинки, куртинки, подушки), что помогает переносить неблагоприятные условия и способствует вегетативному размножению. У одних М. из почек вырастают пластинчатые слоевища, у других - радиально или двусторонне-симметричные побеги. Слоевища и побеги могут быть обоеполыми или однополыми; соответственно М. бывают обоеполыми или разнополыми, однодомными и двудомными. Органы полового размножения - антеридии (См.
Антеридий) и архегонии (См.
Архегоний) - чаще располагаются группами среди многоклеточных стерильных нитей - парафиз и окружены особыми листовидными выростами. В антеридиях (продолговатых мешочках на ножке с однослойной оболочкой) образуются двужгутиковые сперматозоиды, для передвижения которых к яйцеклетке необходима вода. В нижней расширенной части архегония помещается яйцеклетка. Оплодотворение и дальнейшее развитие зиготы происходит в архегонии. Из зиготы за несколько месяцев развивается спорогон - орган бесполого размножения (спороношения), в значительной степени утерявший самостоятельность. Зелёное стеблеобразное тело молодого спорогона затем становится жёлтым, коричневым или тёмно-красным и дифференцируется на верхнюю спороносную часть - коробочку и нижнюю - ножку со стопой, которая врастает в ткань материнского растения. При образовании спор из археспория (спорогенной ткани) происходит
Мейоз. Зигота и спорогон у М. диплоидны и составляют бесполое поколение - спорофит. Протонема, слоевищные и олиственные гаметофоры гаплоидны и относятся к половому поколению - гаметофиту; в основном на них ложатся функции автотрофного питания. Благодаря этому и способности гаметофита к вегетативному размножению цикл развития М. длительное время может происходить без образования спорофита (у некоторых видов спорогоны неизвестны). Гаметофиты М. морфологически разнообразны, однослойные листовидные выросты стебля (филлоиды) отличаются большим варьированием признаков. Спорогоны же во многих группах обнаруживают значительное сходство. На стеблях олиственных М. образуются также хлорофиллоносные листо- или нитевидные выросты - парафиллии, булавовидные волоски, вторичная протонема и нитевидные
Ризоиды, которые, как войлоком, могут обволакивать наземные или подземные части растений. Ризоидами слоевища и стебли прикрепляются к субстрату или плотно соединяются между собой. У М. установлены
Апоспория и
Апогамия, гибридизация и
Полиплоидия, что даёт основания для признания их участия в видообразовательных процессах. Наряду с этим у М. распространено формирование
Клонов, что способствует сохранению местных адаптивных изменений.
Антоцеротовые М. (Anthocerotae) - своеобразная группа из 2 семейств с 6 родами, объединяющими свыше 300 видов, распространённых преимущественно в тропиках, в СССР 4-5 видов. Слоевище у них обычно лопастное или розетковидное с погружёнными в него антеридиями и архегониями. Спорогон, как правило, вытянутый, стручковидный, способный удлиняться благодаря меристеме (См.
Меристема), расположенной в его нижней части, с колонкой в середине и с устьицами в наружных стенках; у основания окружен обёрткой и раскрывается 2 створками сверху вниз. Кроме спор, образуются одно- или многоклеточные бесплодные короткие нити - элатеры, способствующие распространению спор. В клетках слоевища и спорогонов содержится 1 или несколько хроматофоров с
Пиреноидами.
Печёночные М., или печёночники (Hepaticae), подразделяются на 2 подкласса: маршанциевые и юнгерманниевые. В отличие от других М., у печёночников протонема слабо развита, недолговечна и на ней обычно развивается одно слоевищное или листостебельное растение дорзо-вентральной структуры; в клетках гаметофита содержатся одно или несколько масляных телец постоянной формы; ризоиды одноклеточные. Спорогон чаще без хлоропластов, лишён колонки, и на его стенках отсутствуют устьица. У некоторых маршанциевых обнаруживается сходство с антоцеротовыми, поэтому антоцеротовых рассматривали как подкласс печёночных М. Гаметофит печёночников отличается большим разнообразием, спорогоны однотипны. Маршанциевые печёночники (Marchantiidae) - слоевищные формы, обычно в виде дихотомически ветвящихся маленьких розеток или крупных лентовидно стелящихся слоевищ, прикрепляющихся к почве ризоидами; мужские и женские гаметангии погружены в ткань слоевища или приподняты на особых подставках. Преимущественно напочвенные растения, редко скальные или водные, распространены по всему земному шару, особенно богато представлены в тропиках. Известно 16 семейств с 35 родами, объединяющими около 420 видов, включая ископаемую Naiadita из мезозоя. Наиболее примитивны М. из порядка сферокарповых с просто устроенным слоевищем, иногда с масляными тельцами в особых клетках, с однослойной стенкой спорогона и с питательными клетками вместо элатер. Для М. порядка маршанциевых характерно сложное строение слоевища с разделением на основную и ассимиляционную ткань, с масляными тельцами в особых клетках и со склеренхимными волокнами (у некоторых видов). Слоевища часто с воздушными камерами и воздушными устьицами, с нижней стороны с брюшными чешуйками и одноклеточными гладкими и язычковыми ризоидами. Спорогон обычно более сложного строения. Среди них риччиевые рассматриваются как вторично упрощённые, с гаметангиями и спорогоном, развивающимся внутри мелких, чаще розетковидных слоевищ. Юнгерманниевые печёночники (Jungermanniidae) включают группу слоевищных и олиственных растений, в вегетативных клетках которых, как правило, по нескольку масляных телец; ризоиды гладкие, одноклеточные. Эпифитные, напочвенные и наскальные растения особенно богато представлены в субтропиках и тропиках. 45 семейств объединяют свыше 240 родов, включающих около 9 тыс. видов. У представителей порядка метцгериевых слоевище одно- или многослойное, недифференцированное (у некоторых с брюшными или спинными, иногда листовидными чешуйками и стеблями с листовидными лопастями); коробочка двух- или многослойная. Растения порядка гапломитриевых прямостоячие, с трёхрядным листорасположением и коробочками, имеющими однослойные боковые стенки. Наиболее многочисленны и разнообразны М. порядка юнгерманниевых, для которых характерны 2 ряда цельных или лопастных боковых листьев без жилки и более мелкие (иногда отсутствуют) брюшные листья - так называемые амфигастрии; стенки коробочки многослойные.
Лиственные М., или листостебельные М. (Musci), подразделяются на 3 подкласса: сфагновые, андреевые и бриевые. Растения радиально-, редко двусторонне-симметричные, спиральноолиственные; листья цельные, зубчатые или по краю реснитчатые, однослойные, реже дву- или многослойные, с жилкой или без жилки; с многоклеточными, ветвистыми ризоидами. Спорогоны верхушечные или боковые, разнообразные по форме, строению и окраске. Около 15 тыс. видов (по др. данным, до 25 тыс. видов), объединяемых в 660 родов. Сфагновые, или торфяные, М. имеют крупные, прямостоячие стебли с пучковидно расположенными ветвями, на верхушке собранными в головку; беловато-зелёные, жёлтые, бурые или красноватые. Листья однослойные, веточные и стеблевые, без жилки, из чередующихся хлорофиллоносных и водоносных клеток. Спорогон состоит из шаровидной коробочки с крышечкой и недоразвитыми устьицами, без ножки, со стопой, вросшей в безлистную удлинённую архегониальную веточку стебля. Споровый мешок сводообразно покрывает верхнюю часть колонки. Распространены по всему земному шару: преобладают на болотах, в тундре, во влажных лесах. Андреевые М. - мелкие красно-бурые (до чёрных), с однослойными листьями; растут на скалах подушечками. Коробочка удлинённо-яйцевидная, на ложноножке, раскрывается 4-8 створками, без устьиц, со споровым мешком в виде свода над верхней частью колонки. Распространены высоко в горах, а также в северных широтах. Бриевые М. - наиболее обширная, разнообразная группа. Коробочка обычно на настоящей ножке, с устьицами, чаще после опадения крышечки открывается наверху широким отверстием - устьем. У многих по краю устья образуется 1-2 ряда выростов - зубцов (называемых простым или двойным перистомом), которые служат для рассеивания спор. Колонка, как правило, пронизывает споровый мешок. (Цикл развития см.
Лиственные мхи.)
М. распространены повсеместно. Поселяются повсюду, кроме морей, засолённых почв и местообитаний, скрытых под ледниками или подверженных сильной эрозии. Антоцеротовые и печёночные М. распространены в основном в странах с тропическим или умеренным влажным климатом; только немногие приспособились к произрастанию в сухих местах. Лиственные М. растут почти повсеместно, однако наибольшего развития достигают на увлажнённых местах, в лесах, тундре; на болотах М. образуют основную массу торфяных залежей. Интенсивно развиваясь, М. способствуют заболачиванию почв, ухудшают качество лугов и др. с.-х. угодий. В связи с антибиотическими свойствами некоторых М. их применяют иногда как перевязочный материал. Используют в качестве подстилки для скота (преимущественно сфагновые М.), в строительной технике для изготовления плит и др.
Лит.: Флора споровых растений СССР, т. 1, 3, М. - Л., 1952-54; Савич-Любицкая Л. И., Смирнова З. Н., Определитель сфагновых мхов СССР, Л., 1968; их же, Определитель листостебельных мхов СССР. Верхоплодные мхи, Л., 1970; Smith G. M., Cryptogamic botany, 2ed., v. 2, N. Y., 1955; Schuster R. M., The Hepaticae and Anthocerotae of North America, v. 1, N. Y. - L., 1966; Parihar N. S., An introduction to embryophyta, 5 ed., v. 1, Allahabad, 1969.
И. И. Абрамов.
Печёночные мхи: 1 - Plagiochila asplenioides. Листостебельные мхи: 2 - Sphagnum magellanicum, 3 - Sphagnum palustre.
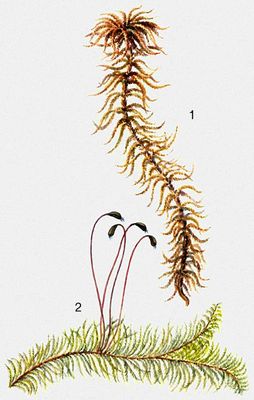
Листостебельные мхи: 1 - Sphagnum fuxum, 2 - Ptilium crista-castrensis.
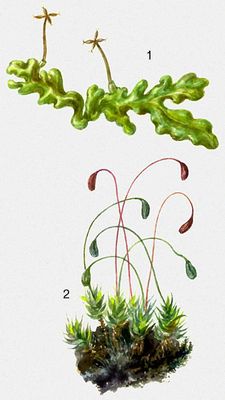
Печёночные мхи: 1 - Riccardia pinguis. Листостебельные мхи: 2 - Funaria hygrometrica.
Печёночные мхи: Conocephalum conicum.
Печёночные мхи: 1 - Marchantia polymorpha. Листостебельные мхи: 2 - Dicranum scoparium, 3 - Sphagnum recurvum, 4 - Splachnum rubrum, 5 - Polytrichum commune, 6 - Fontinalis antipyretica, 7 - Bryum marratii, 8 - Rhytidiadelphus triquetrus.
Атоцеротовые мхи: 1 - Anthoceros punctatus. Печёночные мхи: 2 - Riccia glauca, 3 - Riccia fluitans.

![Культивирование мха в [[биореактор]]е](https://commons.wikimedia.org/wiki/Special:FilePath/Bioreaktor quer2.jpg?width=200 "Культивирование мха в [[биореактор]]е")

![[[Хлоропласт]]ы в клетках ''[[Plagiomnium affine]]'' из семейства ''[[Mniaceae]]''](https://commons.wikimedia.org/wiki/Special:FilePath/Plagiomnium affine laminazellen.jpeg?width=200 "[[Хлоропласт]]ы в клетках ''[[Plagiomnium affine]]'' из семейства ''[[Mniaceae]]''")